
เนื้อหา
- ลักษณะทั่วไปของเซลล์
- โครงสร้างเซลล์ Prokaryotic ทั่วไป
- โครงสร้างของผนังเซลล์แบคทีเรีย
- แบคทีเรียแกรมบวกและแกรมลบ
- ผนังเซลล์แบคทีเรียแกรมบวก
- บทบาทของกรด Teichoic
- ผนังเซลล์แบคทีเรียแกรมลบ
- เครื่องมือของแบคทีเรียแกรมลบ
- ผนังเซลล์ Archaea
- ทำไมผนังเซลล์ถึงสำคัญ?
- ความต้านทานยาปฏิชีวนะ
prokaryotes เป็นตัวแทนหนึ่งในสองการจำแนกประเภทที่สำคัญของชีวิต ส่วนอื่นคือ ยูคาริโอ.
Prokaryotes ถูกแยกจากกันด้วยระดับความซับซ้อนที่ต่ำกว่า พวกเขาทั้งหมดด้วยกล้องจุลทรรศน์แม้ว่าจะไม่ได้เป็นเซลล์เดียว พวกเขาจะแบ่งออกเป็นโดเมน เคีย และ แบคทีเรีย, แต่ส่วนใหญ่ของสายพันธุ์ Prokaryote ที่รู้จักกันดีคือแบคทีเรียซึ่งอยู่บนโลกมาประมาณ 3.5 พันล้านปี
เซลล์โปรคาริโอตไม่มีนิวเคลียสหรือออร์แกเนลล์ที่จับกับเยื่อหุ้มเซลล์ อย่างไรก็ตาม 90 เปอร์เซ็นต์ของแบคทีเรียมีอย่างไรก็ตาม ผนังเซลล์ซึ่งยกเว้นเซลล์พืชและเซลล์เชื้อราบางเซลล์เซลล์ยูคาริโอตขาด ผนังเซลล์เหล่านี้ก่อตัวเป็นชั้นนอกสุดของแบคทีเรียและเป็นส่วนหนึ่งของ แคปซูลแบคทีเรีย.
พวกมันมีความเสถียรและปกป้องเซลล์และมีความสำคัญต่อแบคทีเรียที่สามารถแพร่เชื้อไปยังเซลล์โฮสต์รวมถึงการตอบสนองของแบคทีเรียต่อยาปฏิชีวนะ
ลักษณะทั่วไปของเซลล์
เซลล์ทั้งหมดในธรรมชาติแบ่งปันคุณสมบัติมากมายที่เหมือนกัน หนึ่งในนั้นคือการปรากฏตัวของภายนอก เยื่อหุ้มเซลล์, หรือ เมมเบรนพลาสม่าซึ่งรูปแบบขอบเขตทางกายภาพของเซลล์ในทุกด้าน อีกอย่างคือสารที่เรียกว่า พลาสซึม พบได้ภายในเยื่อหุ้มเซลล์
ที่สามคือการรวมของสารพันธุกรรมในรูปแบบของ ดีเอ็นเอ, หรือ กรด deoxyribonucleic. ที่สี่คือการปรากฏตัวของ ไรโบโซมซึ่งผลิตโปรตีน เซลล์ที่มีชีวิตทุกเซลล์ใช้ ATP (adenosine triphosphate) เป็นพลังงาน
โครงสร้างเซลล์ Prokaryotic ทั่วไป
โครงสร้างของโปรคาริโอตนั้นง่าย ในเซลล์เหล่านี้ DNA แทนที่จะถูกบรรจุภายในนิวเคลียสที่ล้อมรอบภายในเมมเบรนนิวเคลียร์พบว่ารวมตัวกันอย่างหลวม ๆ ในไซโตพลาสซึมในรูปแบบของร่างกายที่เรียกว่า nucleoid.
โดยปกติจะอยู่ในรูปของโครโมโซมแบบวงกลม
ไรโบโซมของเซลล์โปรคาริโอตพบกระจายอยู่ทั่วไซโตพลาสซึมของเซลล์ในขณะที่ยูคาริโอตบางส่วนพบในออร์กาเนลเช่น เครื่อง Golgi และ เอนโดพลาสซึม reticulum. งานของไรโบโซมคือการสังเคราะห์โปรตีน
แบคทีเรียทำซ้ำโดยการแบ่งฟิชชันหรือแบ่งออกเป็นสองส่วนและแบ่งส่วนประกอบของเซลล์อย่างเท่าเทียมกันรวมถึงข้อมูลทางพันธุกรรมในโครโมโซมขนาดเล็กเดี่ยว
การแบ่งเซลล์รูปแบบนี้ไม่จำเป็นต้องใช้ระยะที่แตกต่าง
โครงสร้างของผนังเซลล์แบคทีเรีย
Peptidoglycans ที่ไม่ซ้ำใคร: ผนังเซลล์พืชและผนังเซลล์แบคทีเรียส่วนใหญ่ประกอบด้วยโซ่คาร์โบไฮเดรต
แต่ในขณะที่ผนังเซลล์ของพืชมีเซลลูโลสซึ่งคุณจะเห็นรายการของส่วนผสมของอาหารมากมาย แต่ผนังของเซลล์แบคทีเรียมีสารที่เรียกว่า peptidoglycanซึ่งคุณจะไม่
peptidoglycan นี้ซึ่งก็คือ พบในโปรคาริโอตเท่านั้นมาในรูปแบบต่าง ๆ มันทำให้เซลล์โดยรวมรูปร่างและการป้องกันฟาโรห์สำหรับเซลล์จากการดูถูกทางกล
Peptidoglycans ประกอบด้วยกระดูกสันหลังที่เรียกว่า ไกลแคนซึ่งประกอบด้วยตัวมันเอง กรด muramic และ กลูโคซาซึ่งทั้งสองกลุ่มมีกลุ่มอะเซทิลีนติดอยู่กับอะตอมไนโตรเจน พวกเขายังรวมถึงโซ่เปปไทด์ของกรดอะมิโนที่เชื่อมโยงข้ามกับโซ่เปปไทด์อื่น ๆ ที่อยู่ใกล้เคียง
ความแข็งแรงของปฏิกิริยา "เชื่อมโยง" เหล่านี้แตกต่างกันอย่างมากระหว่าง peptidoglycans ที่แตกต่างกันดังนั้นระหว่างแบคทีเรียที่แตกต่างกัน
ตามที่คุณเห็นจะช่วยให้แบคทีเรียสามารถจำแนกได้เป็นประเภทที่แตกต่างกันโดยขึ้นอยู่กับว่าผนังเซลล์ของพวกเขาทำปฏิกิริยากับสารเคมีอย่างไร
การเชื่อมโยงข้ามเกิดขึ้นจากการกระทำของเอนไซม์ที่เรียกว่า transpeptidaseซึ่งเป็นเป้าหมายของคลาสยาปฏิชีวนะที่ใช้เพื่อต่อสู้กับโรคติดเชื้อในมนุษย์และสิ่งมีชีวิตอื่น ๆ
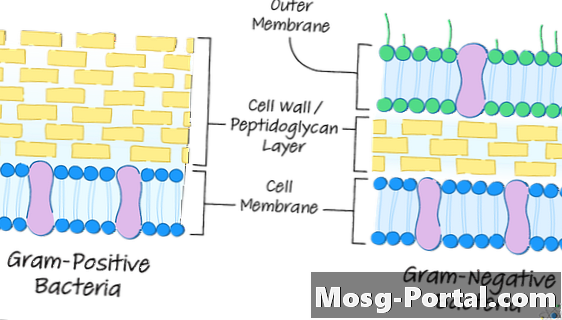
แบคทีเรียแกรมบวกและแกรมลบ
ในขณะที่แบคทีเรียทั้งหมดมีผนังเซลล์องค์ประกอบของมันจะเปลี่ยนจากสปีชีส์หนึ่งไปเป็นสปีชีส์เนื่องจากความแตกต่างในเนื้อหาของเปปทิโดกริแคนที่ผนังเซลล์ส่วนใหญ่หรือส่วนใหญ่ทำ
แบคทีเรียอาจถูกแยกออกเป็นสองประเภทเรียกว่าแกรมบวกและแกรมลบ
เหล่านี้ถูกตั้งชื่อตามนักชีววิทยา ฮันส์คริสเตียนแกรมผู้บุกเบิกชีววิทยาเซลล์ที่พัฒนาเทคนิคการย้อมสีในยุค 1880 เรียกว่า aptly กรัมคราบทำให้แบคทีเรียบางตัวกลายเป็นสีม่วงหรือน้ำเงินและอื่น ๆ กลายเป็นสีแดงหรือสีชมพู
แบคทีเรียชนิดก่อนหน้านี้ถูกเรียกว่า แกรมบวกและคุณสมบัติการย้อมสีนั้นมีสาเหตุมาจากความจริงที่ว่าผนังเซลล์ของพวกมันมีส่วนของ peptidoglycan ที่สูงมากซึ่งสัมพันธ์กับความสมบูรณ์ของผนัง
แบคทีเรียสีแดงหรือชมพูย้อมสีเป็นที่รู้จักกัน แกรมลบและอย่างที่คุณอาจเดาได้แบคทีเรียเหล่านี้มีผนังที่ประกอบด้วย peptidoglycan ในปริมาณเล็กน้อยถึงปานกลาง
ในแบคทีเรียแกรมลบเยื่อบาง ๆ อยู่นอกผนังเซลล์ก่อตัว ซองจดหมายของเซลล์.
ชั้นนี้คล้ายกับพลาสมาเมมเบรนของเซลล์ที่อยู่อีกด้านหนึ่งของผนังเซลล์ใกล้กับด้านในของเซลล์ ในเซลล์แกรมลบบางตัวเช่น อี. โคไลเยื่อหุ้มเซลล์และซองจดหมายนิวเคลียร์สัมผัสกับบางสถานที่จริง ๆ แทรกซึม peptidoglycan ของผนังบาง ๆ
ซองจดหมายนิวเคลียร์นี้ประกอบด้วยโมเลกุลที่ยื่นออกไปด้านนอกที่เรียกว่า lipopolysaccharides, หรือ LPS. ที่ยื่นออกมาจากด้านในของเมมเบรนนี้คือ mopin lipoproteins ที่ติดอยู่ที่ปลายสุดด้านนอกของผนังเซลล์
ผนังเซลล์แบคทีเรียแกรมบวก
แบคทีเรียแกรมบวกมีผนังเซลล์ peptidoglycan หนาประมาณ 20 ถึง 80 นาโนเมตร (นาโนเมตรหรือหนึ่งในพันล้านส่วนของหนึ่งเมตร)
ตัวอย่าง ได้แก่ Staphylococci, Streptococci, แลคโตบาซิลลัส และ บาซิลลัส สายพันธุ์
แบคทีเรียเหล่านี้เปื้อน สีม่วงหรือสีแดงแต่โดยปกติแล้วจะเป็นสีม่วงโดยมีคราบ Gram เนื่องจาก peptidoglycan ยังคงใช้สีย้อมสีม่วงที่ใช้ในขั้นตอนแรกเมื่อการเตรียมการถูกล้างด้วยแอลกอฮอล์ในภายหลัง
ผนังเซลล์ที่แข็งแรงกว่านี้มีแบคทีเรียแกรมบวกที่ป้องกันได้มากขึ้นจากการดูถูกนอกส่วนใหญ่เมื่อเทียบกับแบคทีเรียแกรมลบแม้ว่า เนื้อหา peptidoglycan สูง ของสิ่งมีชีวิตเหล่านี้ทำให้กำแพงของพวกเขาเป็นป้อมปราการแบบหนึ่งมิติทำให้กลายเป็นกลยุทธ์ที่ง่ายกว่าในการทำลายมัน
••• Sciencingโดยทั่วไปแล้วแบคทีเรียแกรมบวกมักไวต่อยาปฏิชีวนะที่กำหนดเป้าหมายที่ผนังเซลล์มากกว่าที่จะเป็นสายพันธุ์แกรมลบเนื่องจากสัมผัสกับสภาพแวดล้อมซึ่งต่างจากการนั่งอยู่ข้างใต้หรือภายในซองจดหมายของเซลล์
บทบาทของกรด Teichoic
ชั้น peptidoglycan ของแบคทีเรียแกรมบวกมักจะมีโมเลกุลสูงที่เรียกว่า กรด Teichoic, หรือ ครู.
เหล่านี้เป็นโซ่คาร์โบไฮเดรตที่เข้าถึงและบางครั้งก็ผ่านชั้น peptidoglycan
เชื่อว่า TA จะทำให้ peptidoglycan ที่อยู่รอบตัวมันคงที่โดยการทำให้มันมีความแข็งมากกว่าโดยการออกแรงทางเคมี
TA มีส่วนรับผิดชอบต่อความสามารถของแบคทีเรียแกรมบวกบางชนิดเช่นสเตรปโตคอกคคัสในการจับกับโปรตีนจำเพาะบนพื้นผิวของเซลล์เจ้าบ้านซึ่งอำนวยความสะดวกในการติดเชื้อและในหลายกรณี
เมื่อแบคทีเรียหรือจุลินทรีย์อื่น ๆ มีความสามารถในการก่อให้เกิดโรคติดเชื้อพวกเขาจะเรียกว่า เป็นเชื้อโรค.
ผนังเซลล์ของแบคทีเรียของ ครอบครัวมัยโคแบคทีเรียนอกเหนือจากการมี peptidoglycan และ TAs แล้วยังมีเลเยอร์“ ข้าวเหนียว” ที่ทำจาก กรด mycolic. แบคทีเรียเหล่านี้รู้จักกันในนาม“กรดอย่างรวดเร็ว” เพราะคราบประเภทนี้มีความจำเป็นในการเจาะเลเยอร์ขี้ผึ้งเพื่อให้สามารถตรวจด้วยกล้องจุลทรรศน์ได้
ผนังเซลล์แบคทีเรียแกรมลบ
แบคทีเรียแกรมลบเช่นคู่ของแกรมบวกมีผนังเซลล์ peptidoglycan
อย่างไรก็ตามผนังนั้นบางมากเพียง 5 ถึง 10 นาโนเมตรหนา ผนังเหล่านี้ไม่เปื้อนสีม่วงกับสีแกรมเพราะเนื้อหาที่มีขนาดเล็กกว่าของ peptidoglycan หมายความว่าผนังไม่สามารถเก็บสีย้อมได้มากเมื่อการเตรียมการถูกล้างด้วยแอลกอฮอล์ทำให้เกิดสีชมพูหรือสีแดงในที่สุด
ตามที่ระบุไว้ข้างต้นผนังเซลล์ไม่ได้อยู่ด้านนอกสุดของแบคทีเรียเหล่านี้ แต่ถูกปกคลุมด้วยเมมเบรนพลาสม่าอื่น, ซองเซลล์หรือเมมเบรนด้านนอก
ชั้นนี้มีความหนา 7.5 ถึง 10 นาโนเมตรหนาหรือหนากว่าผนังเซลล์
ในแบคทีเรียแกรมลบส่วนใหญ่ซองเซลล์จะเชื่อมโยงกับโมเลกุลไลโปโปรตีนชนิดหนึ่งที่เรียกว่า Brauns lipoprotein ซึ่งในทางกลับกันจะเชื่อมโยงกับ peptidoglycan ของผนังเซลล์
เครื่องมือของแบคทีเรียแกรมลบ
โดยทั่วไปแล้วแบคทีเรียแกรมลบมักมีความไวต่อยาปฏิชีวนะน้อยกว่าที่กำหนดไว้ที่ผนังเซลล์เนื่องจากไม่ได้สัมผัสกับสิ่งแวดล้อม มันยังมีเมมเบรนด้านนอกสำหรับการป้องกัน
นอกจากนี้ในแบคทีเรียแกรมลบเมทริกซ์ที่มีลักษณะคล้ายเจลจะใช้พื้นที่ภายในผนังเซลล์และนอกเมมเบรนพลาสมาที่เรียกว่าพื้นที่ periplasmic
ส่วนประกอบ peptidoglycan ของผนังเซลล์ของแบคทีเรียแกรมลบมีความหนาเพียง 4 นาโนเมตร
ในกรณีที่ผนังเซลล์แบคทีเรียแกรมบวกจะมี peptidoglycans มากขึ้นในการให้สารผนังข้อผิดพลาดแกรมลบมีเครื่องมืออื่น ๆ ที่อยู่ในเยื่อหุ้มชั้นนอก
โมเลกุล LPS แต่ละอันประกอบไปด้วยลิพิดย่อยที่อุดมด้วยกรดไขมัน A, โพลีแซคคาไรด์แกนกลางขนาดเล็กและสายโซ่ O-side ทำจากโมเลกุลคล้ายน้ำตาล สายโซ่ O-side นี้ก่อให้เกิดด้านนอกของ LPS
องค์ประกอบที่แน่นอนของโซ่ด้านข้างนั้นแตกต่างกันไปตามชนิดของแบคทีเรีย
ส่วนของสายโซ่ O-side ที่รู้จักกันเป็นแอนติเจนสามารถระบุได้ผ่านการทดสอบในห้องปฏิบัติการเพื่อระบุสายพันธุ์แบคทีเรียที่ทำให้เกิดโรคเฉพาะ (“ ความเครียด” เป็นสายพันธุ์ย่อยของสายพันธุ์แบคทีเรียเช่นสายพันธุ์ของสุนัข)
ผนังเซลล์ Archaea
เคีย มีความหลากหลายมากกว่าแบคทีเรียและผนังเซลล์ของพวกมัน โดยเฉพาะอย่างยิ่งผนังเหล่านี้ไม่มี peptidoglycan
แต่มักจะมีโมเลกุลที่เรียกว่าคล้ายกัน pseudopeptidoglycan, หรือ pseudomurein. ในสารนี้ส่วนหนึ่งของ peptidoglycan ปกติที่เรียกว่า NAM จะถูกแทนที่ด้วยหน่วยย่อยที่แตกต่างกัน
อาร์เคียบางคนอาจมีชั้นของ ไกลโคโปรตีน หรือ polysaccharides แทนที่ผนังเซลล์แทน pseudopeptidoglycan ในที่สุดเช่นเดียวกับแบคทีเรียบางชนิดอาร์เคียบางอันก็หายไปจากผนังเซลล์โดยสิ้นเชิง
Archaea ที่มี pseudomurein ได้แก่ ไม่ไวต่อยาปฏิชีวนะในกลุ่มยาเพนิซิลิน เพราะยาเหล่านี้เป็นตัวยับยั้ง transpeptidase ที่ทำหน้าที่ขัดขวางการสังเคราะห์ peptidoglycan
ใน Archaea เหล่านี้ไม่มีการสังเคราะห์ peptidoglycans และดังนั้นจึงไม่มีอะไรที่เพนิซิลินจะทำ
ทำไมผนังเซลล์ถึงสำคัญ?
เซลล์แบคทีเรียที่ไม่มีผนังเซลล์อาจมีโครงสร้างพื้นผิวของเซลล์เพิ่มเติมนอกเหนือจากที่กล่าวถึงเช่น glycocalyces เอกพจน์คือ glycocalyx) และ S-layer
glycocalyx เป็นเสื้อคลุมของโมเลกุลคล้ายน้ำตาลที่มาในสองประเภทหลัก: แคปซูล และ ชั้นน้ำเมือก. แคปซูลเป็นชั้นของโพลีแซคคาไรด์หรือโปรตีนที่มีการจัดระเบียบอย่างดี ชั้นเมือกนั้นมีการจัดเรียงอย่างแน่นหนาน้อยกว่าและมันติดแน่นกับผนังเซลล์ด้านล่างน้อยกว่า glycocalyx
เป็นผลให้ glycocalyx ทนต่อการถูกชะล้างออกไปได้มากกว่าในขณะที่ชั้นเมือกสามารถถูกแทนที่ได้ง่ายขึ้น ชั้นเมือกอาจประกอบด้วยโพลีแซคคาไรด์, ไกลโคโปรตีนหรือไกลคอลิพิด
รูปแบบทางกายวิภาคเหล่านี้ให้ความสำคัญกับคลินิกอย่างมาก
Glycocalyces ช่วยให้เซลล์สามารถยึดติดกับพื้นผิวบางอย่างช่วยในการสร้างอาณานิคมของสิ่งมีชีวิตที่เรียกว่า ไบโอฟิล์ม ที่สามารถสร้างหลายเลเยอร์และปกป้องบุคคลในกลุ่ม ด้วยเหตุนี้แบคทีเรียส่วนใหญ่ในป่าจึงอาศัยอยู่ในแผ่นชีวะที่เกิดจากชุมชนแบคทีเรียผสม แผ่นชีวะขัดขวางการกระทำของยาปฏิชีวนะเช่นเดียวกับยาฆ่าเชื้อ
คุณลักษณะทั้งหมดเหล่านี้มีส่วนทำให้ความยากลำบากในการกำจัดหรือลดจุลินทรีย์และกำจัดการติดเชื้อ
ความต้านทานยาปฏิชีวนะ
แบคทีเรียสายพันธุ์ที่มีความต้านทานตามธรรมชาติต่อยาปฏิชีวนะที่ได้รับจากการกลายพันธุ์ที่ได้เปรียบคือ "เลือก" ในประชากรมนุษย์เพราะสิ่งเหล่านี้เป็นแมลงที่ถูกทิ้งไว้ข้างหลังเมื่อยาปฏิชีวนะที่ไวต่อเชื้อถูกฆ่าตายและ "superbugs" ทำให้เกิดโรค
ภายในทศวรรษที่สองของศตวรรษที่ 21 แบคทีเรียชนิดกรัมลบได้กลายเป็นดื้อต่อยาปฏิชีวนะมากขึ้นซึ่งนำไปสู่การเจ็บป่วยและเสียชีวิตที่เพิ่มขึ้นจากการติดเชื้อและผลักดันค่าใช้จ่ายด้านการดูแลสุขภาพ ความต้านทานยาปฏิชีวนะเป็นตัวอย่างตามแบบฉบับของส่วนตามธรรมชาติในช่วงเวลาที่สังเกตได้กับมนุษย์
ตัวอย่างรวมถึง:
นักวิจัยด้านการแพทย์กำลังทำงานเพื่อให้ทันกับโรคจิตดื้อยาในจำนวนที่มีต่อการแข่งขันทางจุลชีววิทยา