เนื้อหา
- ข้อมูลพื้นฐานเกี่ยวกับกลูโคส
- เส้นทาง Glycolysis ที่สมบูรณ์
- Glycolysis: ระยะการลงทุน
- Glycolysis: Return phase
- ระเบียบของ Glycolysis
- Glycolysis Enzyme Activation
- Glycolysis Enzyme Inhibition
- เพิ่มเติมเกี่ยวกับกฎระเบียบ Hexokinase
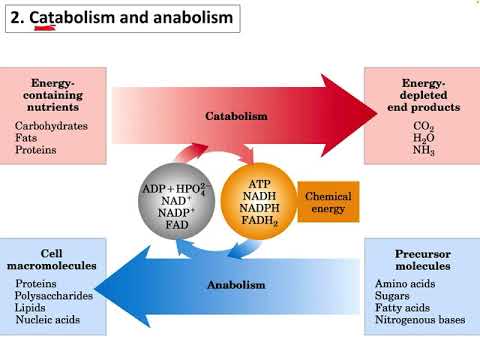
glycolysis เป็นกระบวนการเผาผลาญสากลในหมู่สิ่งมีชีวิตในโลก ชุด 10 ปฏิกิริยาในโปรโตปลาสซึมของเซลล์ทั้งหมดแปลงโมเลกุลของน้ำตาลหกคาร์บอน กลูโคส เป็นสองโมเลกุลของไพรูเวต, สองโมเลกุลของ ATP และสองโมเลกุลของ NADH
เรียนรู้เกี่ยวกับ glycolysis
ใน prokaryotesซึ่งเป็นสิ่งมีชีวิตที่ง่ายที่สุด glycolysis เป็นเกมการเผาผลาญเซลลูล่าร์เดียวในเมือง สิ่งมีชีวิตเหล่านี้เกือบทั้งหมดประกอบด้วยเซลล์เดียวที่มีเนื้อหาค่อนข้างน้อยมีความต้องการเมแทบอลิซึมที่ จำกัด และ glycolysis ก็เพียงพอที่จะให้พวกมันเจริญเติบโตและสืบพันธุ์ในกรณีที่ไม่มีปัจจัยแข่งขัน ยูคาริโอในขณะที่ไกลคอลออกมาเป็นอาหารจานที่ต้องการก่อนอาหารจานหลักของการหายใจแบบแอโรบิค
การสนทนาของ glycolysis มักมุ่งเน้นไปที่เงื่อนไขที่ชอบเช่นพื้นผิวที่เพียงพอและความเข้มข้นของเอนไซม์ มีการกล่าวถึงน้อย แต่สิ่งสำคัญก็คือสิ่งที่อาจเกิดขึ้นจากการออกแบบ ยับยั้ง อัตรา glycolysis แม้ว่าเซลล์ต้องการพลังงาน แต่การใช้วัตถุดิบอย่างต่อเนื่องผ่านโรงสีไกลคอลไลซิสนั้นไม่ได้เป็นผลที่ต้องการเสมอไป โชคดีสำหรับเซลล์ผู้เข้าร่วมจำนวนมากใน glycolysis มีความสามารถในการส่งผลกระทบต่อความเร็ว
ข้อมูลพื้นฐานเกี่ยวกับกลูโคส
กลูโคสเป็นน้ำตาลหกคาร์บอนที่มีสูตร C6H12O6. (Fun biomolecule trivia: คาร์โบไฮเดรตทุกชนิดไม่ว่าจะเป็นน้ำตาลแป้งหรือเส้นใยที่ไม่ละลายน้ำมีสูตรทางเคมีทั่วไป Cยังไม่มีข้อความH2NOยังไม่มีข้อความ.) มีมวลโมลาร์ 180 กรัมคล้ายกับกรดอะมิโนที่หนักกว่าในแง่ของขนาด มันสามารถแพร่กระจายเข้าและออกจากเซลล์ผ่านพลาสมาเมมเบรนได้อย่างอิสระ
กลูโคสเป็น monosaccharide ซึ่งหมายความว่ามันไม่ได้เกิดจากการรวมน้ำตาลที่มีขนาดเล็กลง ฟรักโทสเป็น monosaccharide ในขณะที่ซูโครส ("น้ำตาลตาราง") เป็นไดแซ็กคาไรด์ที่ประกอบจากโมเลกุลกลูโคสและโมเลกุลฟรุกโตส
ยวดกลูโคสอยู่ในรูปของวงแหวนซึ่งแสดงเป็นรูปหกเหลี่ยมในไดอะแกรมส่วนใหญ่ อะตอมของแหวนห้าในหกตัวนั้นเป็นกลูโคสในขณะที่อะตอมที่หกก็คือออกซิเจน คาร์บอน -6 อยู่ในเมธิล (- CH)3) กลุ่มนอกวงแหวน
เส้นทาง Glycolysis ที่สมบูรณ์
สูตรสมบูรณ์สำหรับผลรวมของ 10 ปฏิกิริยาของ glycolysis คือ:
C6H12O6 + 2 NAD+ + 2 Pi + 2 ADP → 2 CH3(C = O) COOH + 2 ATP + 2 NADH + 2 H+
กล่าวคือโมเลกุลของกลูโคสจะถูกเปลี่ยนเป็นโมเลกุลสองกลูโคสซึ่งจะสร้าง 2 ATP และ 2 NADH (รูปแบบที่ลดลงของ nicotinamide adenine dinucleotide ซึ่งเป็น "ผู้ให้บริการอิเล็กตรอน" ทั่วไปในชีวเคมี)
โปรดทราบว่าไม่จำเป็นต้องใช้ออกซิเจน แม้ว่าไพรูเวตจะมีการบริโภคแอโรบิกในขั้นตอนของการหายใจอย่างสม่ำเสมอ แต่ glycolysis เกิดขึ้นในสิ่งมีชีวิตแบบแอโรบิกและแบบไม่ใช้ออกซิเจนเหมือนกัน
Glycolysis: ระยะการลงทุน
Glycolysis แบ่งออกเป็นสองส่วนคลาสสิก: "ขั้นตอนการลงทุน" ซึ่งต้องใช้ 2 ATP (adenosine triphosphate, "พลังงานสกุลเงิน" ของเซลล์) เพื่อรูปร่างโมเลกุลกลูโคสเป็นสิ่งที่มีศักยภาพพลังงานและ "ผลตอบแทน" หรือ "การเก็บเกี่ยว" ระยะที่ 4 ATP ถูกสร้างขึ้นผ่านการแปลงของโมเลกุลสามคาร์บอนหนึ่งอัน (glyceraldehyde-3-phosphate หรือ GAP) ไปเป็นไพรูเวต ซึ่งหมายความว่า ATP จะถูกสร้างขึ้นทั้งหมด 4 -2 = 2 ATP ต่อโมเลกุลกลูโคส
เมื่อกลูโคสเข้าสู่เซลล์จะเป็นฟอสโฟรีเลต (เช่นมีกลุ่มฟอสเฟตติดอยู่) ภายใต้การทำงานของเอนไซม์ hexokinase. เอนไซม์นี้หรือตัวเร่งปฏิกิริยาโปรตีนเป็นหนึ่งในเอนไซม์ที่สำคัญที่สุดในการควบคุม glycolysis 10 ปฏิกิริยาใน glycolysis แต่ละครั้งจะถูกเร่งโดยเอนไซม์หนึ่งเอนไซม์และเอนไซม์นั้นจะเร่งปฏิกิริยาเพียงปฏิกิริยาเดียวเท่านั้น
กลูโคส -6- ฟอสเฟต (G6P) ซึ่งเป็นผลมาจากขั้นตอน phosphorylation นี้จะถูกแปลงเป็นฟรักโทส -6- ฟอสเฟต (F6P) ก่อนที่จะเกิดการเรืองแสงที่สองครั้งนี้ในทิศทางของ phosphofructokinaseอีกเอนไซม์ที่สำคัญ ส่งผลให้เกิดการสร้างฟรักโทส -16-bisphosphate (FBP) และระยะแรกของ glycolysis เสร็จสมบูรณ์
Glycolysis: Return phase
Fructose-1,6-bisphosphate ถูกแบ่งออกเป็นคู่ของโมเลกุลสามคาร์บอน, dihydroxyacetone phosphate (DHAP) และ glyceraldehyde-3-phosphate (GAP) DHAP จะถูกแปลงเป็น GAP อย่างรวดเร็วดังนั้นผลกระทบสุทธิของการแยกคือการสร้างโมเลกุลสามคาร์บอนสองตัวที่เหมือนกันจากโมเลกุลคาร์บอนเดียวหกตัว
GAP จะถูกแปลงโดยเอนไซม์ glyceraldehyde-3-phosphate dehydrogenase เป็น 1,3-diphosphoglycerate นี่เป็นขั้นตอนที่ยุ่ง NAD+ ถูกแปลงเป็น NADH และ H+ ใช้ไฮโดรเจนอะตอมที่หลุดออกจาก GAP แล้วโมเลกุลก็จะถูกฟอสโฟริ
ในขั้นตอนที่เหลือซึ่งเปลี่ยน 1,3-diphosphoglycerate เป็น pyruvate ฟอสเฟตทั้งสองจะถูกลบออกตามลำดับจากโมเลกุลสามคาร์บอนเพื่อสร้าง ATP เนื่องจากทุกอย่างหลังจากการแยก FBP เกิดขึ้นสองครั้งต่อโมเลกุลกลูโคสนั่นหมายความว่า 2 NADH, 2 H+ และ 4 ATP ถูกสร้างขึ้นในเฟสส่งคืนสำหรับสุทธิ 2 NADH, 2 H+ และ 2 ATP
อ่านเพิ่มเติมเกี่ยวกับผลลัพธ์สุดท้ายของ glycolysis
ระเบียบของ Glycolysis
เอนไซม์สามตัวที่มีส่วนร่วมใน glycolysis มีบทบาทสำคัญในการควบคุมกระบวนการ มีการกล่าวถึงสอง hexokinase และ phosphofructokinase (หรือ PFK) แล้ว ที่สาม, ไคเนส pyruvateมีหน้าที่ในการเร่งปฏิกิริยา glycolysis สุดท้ายการเปลี่ยน phosphoenolpyruvate (PEP) เป็น pyruvate
เอนไซม์แต่ละตัวมี activators เช่นกัน ยับยั้ง. หากคุณคุ้นเคยกับวิชาเคมีและแนวคิดของการยับยั้งข้อเสนอแนะคุณอาจสามารถคาดการณ์เงื่อนไขที่ทำให้เอนไซม์ที่ได้รับนั้นเร่งความเร็วหรือทำให้กิจกรรมช้าลง ตัวอย่างเช่นหากบริเวณหนึ่งของเซลล์อุดมไปด้วย G6P คุณคาดหวังว่าเฮกซาไคเนสจะค้นหาโมเลกุลกลูโคสที่หลงเหลืออยู่อย่างจริงจังหรือไม่? คุณอาจจะไม่ได้เพราะภายใต้เงื่อนไขเหล่านี้ไม่จำเป็นต้องสร้าง G6P เพิ่มเติมอย่างเร่งด่วน และคุณจะถูกต้อง
Glycolysis Enzyme Activation
ในขณะที่ hexokinase ถูกยับยั้งโดย G6P มันถูกเปิดใช้งานโดย AMP (adenosine monophosphate) และ ADP (adenosine diphosphate) เช่นเดียวกับ PFK และ pyruvate kinase นี่เป็นเพราะระดับที่สูงขึ้นของ AMP และ ADP โดยทั่วไปหมายถึงระดับที่ต่ำกว่าของ ATP และเมื่อ ATP อยู่ในระดับต่ำแรงผลักดันของ glycolysis ที่จะเกิดขึ้นนั้นสูง
kinase Pyruvate ยังเปิดใช้งานโดย fructose-1,6-bisphosphate ซึ่งทำให้รู้สึกเพราะ FBP มากเกินไปแสดงว่า glycolysis กลางสะสมต้นน้ำและสิ่งที่ต้องเกิดขึ้นเร็วขึ้นที่ปลายหางของกระบวนการ นอกจากนี้ฟรุกโตส -2,6-bisphosphate ยังเป็นสารกระตุ้นของ PFK
Glycolysis Enzyme Inhibition
Hexokinase ดังที่กล่าวไว้ถูกยับยั้งโดย G6P PFK และ pyruvate kinase ทั้งคู่ถูกยับยั้งโดยการมีอยู่ของ ATP ด้วยเหตุผลพื้นฐานเดียวกันกับที่พวกมันถูกกระตุ้นโดย AMP และ ADP: สถานะพลังงานของเซลล์จะช่วยลดอัตรา glycolysis
PFK ยังถูกยับยั้งโดย ซิทริทส่วนประกอบของวงจร Krebs ที่เกิดขึ้นจากการหายใจแบบแอโรบิค Pyruvate kinase ถูกยับยั้งโดย acetyl CoAซึ่งเป็นโมเลกุลที่ไพรูเวตถูกเปลี่ยนเป็นหลังจาก glycolysis สิ้นสุดและก่อนที่วงจร Krebs จะเริ่มต้น (อันที่จริง acetyl CoA รวมกับ oxaloacetate ในขั้นตอนแรกของวงจรเพื่อสร้างซิเตรต) ในที่สุดกรดอะมิโนอะลานีนก็ยังยับยั้งเอนไซม์ไพรูเวตไคเนส
เพิ่มเติมเกี่ยวกับกฎระเบียบ Hexokinase
คุณอาจคาดหวังว่าผลิตภัณฑ์อื่น ๆ ของ glycolysis นอกเหนือจาก G6P ในการยับยั้ง hexokinase เนื่องจากการปรากฏตัวของพวกเขาในปริมาณที่สำคัญดูเหมือนจะบ่งบอกถึงความต้องการ G6P ที่ลดลง อย่างไรก็ตามมีเพียง G6P เท่านั้นที่ยับยั้ง hexokinase ทำไมนี้
เหตุผลนั้นค่อนข้างง่าย: จำเป็นต้องใช้ G6P สำหรับเส้นทางการเกิดปฏิกิริยานอกเหนือจาก glycolysis, รวมถึง pentose phosphate shunt และ การสังเคราะห์ไกลโคเจน. ดังนั้นหากโมเลกุลปลายน้ำที่นอกเหนือจาก G6P สามารถยับยั้ง hexokinase จากการทำงานของมันเส้นทางการเกิดปฏิกิริยาอื่น ๆ เหล่านี้ก็จะช้าลงหากขาด G6P ที่เข้าสู่กระบวนการและดังนั้นจึงเป็นตัวแทนของความเสียหายของการเรียงลำดับ